<p>Rosenbaum, J.C., and A.E. Carlson (2024)&n
Rosenbaum, J.C., and A.E. Carlson (2024) The SARS coronavirus accessory protein ORF3a rescues potassium conductance in yeast. micropublication Biology 10.17912/micropub.biology.001129

Ion Channels and Fertilization
Our lab using biophysical tools to study excitable cells. We currently have two research directions, uncovering the earliest events of fertilization, and studying how ion channels are regulated.
Fertilization: Fertilization of an egg by more than one sperm presents one of the earliest barriers to successful reproduction. Fertilization by multiple sperm, a condition known as polyspermy, leads to gross chromosomal abnormalities and is lethal to the developing embryos. Due to these catastrophic consequences, eggs employ multiple processes that protect the protect the nascent embryo. The two most widely studied are called the fast and slow blocks to polyspermy. As their names imply, these blocks occur at different times to fertilization. In the slow block to polyspermy, eggs release material into the extracellular space, and within minutes of its release, this material transforms the egg from a gamete ready for sperm entry, to a structure unable to permit fertilization. The slow block is used by eggs from nearly all sexually reproducing animals. By contrast, the fast block to polyspermy is mainly used by eggs from external fertilizers.
![]()
In normal fertilization, the embryonic cleavage furrows develop symmetrically and transverse the entire embryo (center photo at 8 cell stage). By contrast, the cleavage furrows develop irregularly following fertilization by more than one sperm (right photo) (Wozniak, Mayfield, & Carlson, unpublished).
In the fast block, fertilization activates a depolarization of the egg. By currently unknown mechanisms, sperm can bind to, but will not enter, a depolarized egg. Although fertilization-signaled depolarizations have been recorded for many years, the signaling pathways that activate the fast block, including the molecular identity of the channel that conducts the depolarizing currents, was not yet known for any species.
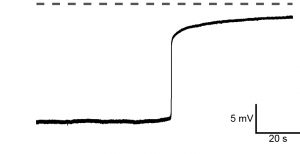
Fertilization-evoked depolarization in X. laevis eggs (recording by Wozniak, unpublished). Dashed ine denotes 0 mV.
The channels that mediate this depolarization and the signaling pathways that regulate it were unknown for any species. We recently uncovered that in eggs from the African clawed frog, Xenopus laevis, fertilization opens the Ca2+-activated Cl- channel TMEM16A. Opening of TMEM16A allows the anion Cl- to leave the egg, and thereby makes the charge of the membrane more positive. To open TMEM16A channels, fertilization activates a phospholipase C (PLC), which cleaves the membrane anchored lipid PIP2 to make the soluble signaling molecule IP3. This IP3 than activates the IP3 receptor to allow Ca2+ to be released from the ER and open TMEM16A channels.

Our established signaling pathway for how fertilization activates the fast block to polyspermy in X. laevis eggs.
We now seek to uncover how fertilization kicks off this pathway. To do so, we use various techniques including electrophysiology and fluorescence imaging to establish the timeline of events.
Ion channel regulation: In addition to playing an important role in X. laevis fertilization, the Ca2+-activated Cl- channel TMEM16A plays an important role in human health. TMEM16A channels play important physiologic roles in diverse tissues including excitability of the insulin-producing pancreatic β cells and excretion by the salivary gland. Overexpression of TMEM16A also plays an important role in the development of several diseases. For example, 80% of craniofacial squamous cell carcinomas are marked by TMEM16A overexpression, and TMEM16a inhibition slows the progression of this cancer. TMEM16A channel expression is increased in airway smooth muscle and secretory cells of asthmatic patients, and blocking its currents attenuates the major symptoms of the disease by restoring cholinergic responsiveness and reducing mucus secretion. Recently, TMEM16A has been proposed to be a novel target for cystic fibrosis treatments. Despite this robust body of evidence documenting a critical role for TMEM16A in human health and disease, little is known about how this channel is regulated in vivo; it is imperative to understand how this channel is regulated in order to exploit this regulation therapeutically.
We have uncovered that the ability of TMEM16A channels to pass Cl- ions depends on the membrane phospholipid PIP2. Using a diverse set of biophysical tools, we now seek to uncover how PIP2 regulates the ability of TMEM16A channels to open and close.
Dr. Carlson recieved the Paul F. Cranefield Award from the Society of General Physiologists in 2019.